



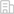

DNA N6-甲基腺嘌呤(N6-methyladenine, 6mA)已被報道參與真核生物的轉錄調控等關鍵生物學過程 [1-6]。然而,6mA與轉錄的關系在不同真核生物中表現出明顯差異,其參與轉錄調控的具體機制尚未充分闡明。嗜熱四膜蟲(Tetrahymena thermophila)作為一種重要的單細胞真核模式生物,已被報道含有較高的6mA水平,由甲基化酶AMT1(Adenine Methyltran sferase 1)催化,且6mA與轉錄存在正相關關系,是研究6mA與轉錄調控機制的理想體系。
2025年1月,中國海洋大學海洋生物多樣性與進化研究所高珊課題組在Nucleic Acids Research(《核酸研究》)雜志發表題為“Methyl-dependent auto-regulation of the DNA N6-adenine methyltransferase AMT1 in the unicellular eukaryote Tetrahymena thermophila”(單細胞真核生物嗜熱四膜蟲DNA N6-腺嘌呤甲基化酶AMT1的自調控)的研究成果。這項研究以四膜蟲為模式材料,發現了6mA甲基化酶AMT1的自調控和轉錄調控機制,并且探究了其在接合生殖時期的動態分布及生理功能,為6mA作為表觀遺傳標記在真核生物中參與轉錄調控和其他生理功能調控提供了重要線索,為未來進一步探索真核生物中6mA的功能調控機制奠定了理論基礎。

團隊前期以四膜蟲為研究材料,鑒定出AMT1是四膜蟲的主要6mA甲基化酶 [7]。AMT1基因缺失會造成細胞生長緩慢,且伸縮泡異常增大。然而,AMT1如何參與基因表達調控,從而影響細胞生長發育的分子機制仍未知。
該工作發現AMT1關鍵酶活位點突變后不僅導致全基因組6mA水平的顯著下降(圖1A-C),自身基因的mRNA水平和蛋白水平也顯著下降(圖1D-E)。通過對AMT1基因的染色質環境進行分析發現,其具有典型的6mA分布偏好性特征,包括:穩定的核小體占位,高水平的H3K4me3和H2A.Z(圖1F),以及相對較高的6mA水平(圖1G)。基于此,推測AMT1基因自身的轉錄水平可能受6mA調控。

圖1. AMT1關鍵酶活位點突變后AMT1自身的mRNA水平,蛋白水平以及表觀遺傳標記的分布情況。(A)AMT1的基因和蛋白模型。(B)AMT1突變后的6mA 免疫熒光染色結果。(C)AMT1突變后的6mA質譜結果。(D)AMT1突變后AMT1(HA)的蛋白印跡結果。(E)AMT1突變后AMT1基因的mRNA表達水平。(F)AMT1基因的6mA、H2A.Z、H3K4me3及核小體分布。(G)所有基因的6mA水平。
進一步地,該工作對AMT1突變株系進行了單堿基分辨率的SMRT-CCS測序。結果發現, AMT1突變后6mA水平顯著下降(圖2A-B),與上述6mA免疫熒光染色和質譜結果一致。更為重要的是,AMT1基因上的6mA水平明顯降低(圖2D-F)。轉錄組測序也顯示,AMT1基因的mRNA表達水平隨之下降(圖2D)。上述結果表明AMT1可能是通過其催化活性調控自身基因的6mA水平,進一步影響其轉錄水平。同時,該工作利用異位過表達野生型AMT1蛋白的拯救實驗,發現野生型AMT1的回補可以回升AMT1基因的6mA水平和mRNA水平(圖2G)。上述結果證實了AMT1催化的6mA可調控自身的轉錄水平,形成了一個正反饋的轉錄激活機制。

圖2. AMT1突變和敲降后的6mA水平分析。(A)6mA甲基化模式。(B)AMT1突變和敲降后的6mA甲基化模式。(C)AMT1突變和敲降后的整體6mA分布情況。(D)AMT1基因的6mA分布和mRNA表達情況。(E)AMT1突變后AMT1基因的6mA 單分子模式。(F)AMT1突變和敲降后所有基因的6mA水平變化。(G)拯救細胞中AMT1基因的6mA和mRNA表達水平。
基于6mA與轉錄存在正相關關系,該工作還整合分析了AMT1敲除、突變、敲降三種細胞的SMRT-CCS和轉錄組數據,鑒定出141個高置信的AMT1-6mA依賴的基因。這些基因主要參與調控氧化還原酶活性,代謝以及跨膜轉運等通路,解釋了AMT1損傷后導致的生長減緩以及伸縮泡異常增大等表型。這些發現為6mA參與轉錄調控提供了更多證據,表明6mA可能作為一種轉錄激活的表觀遺傳標記,參與細胞內生理過程的調控(圖3)。

圖3. AMT1自調控和轉錄調控的模式圖。
該研究由中國海洋大學海洋生物多樣性與進化研究所高珊課題組完成。高珊課題組博士生畢業生段麗麗和博士生李海程為該文章的共同第一作者。王媛媛副教授和高珊教授為文章的通訊作者。日本基礎研究所的Kensuke Kataoka助理教授,中國海洋大學醫藥學院宋妮老師和進化所馬洪鋼老師,高珊課題組碩士生鞠艾利、博士生張喆、牛俊驊、張雨苗、刁靜涵和博士后劉永強對本文亦有重要貢獻。